研究用語辞典
イントロンとエキソンとは
生物学系
最終更新日:2023.09.14
シェアする:
概要
真核生物では、タンパク質の情報に相当する部分が遺伝子DNA中で分断されている場合がほとんどです。遺伝情報がコードされている部分をエキソン(翻訳配列)といい、遺伝情報がコードされていない部分をイントロン(非翻訳、介在配列)といいます。
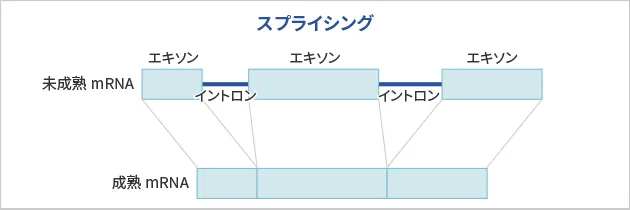
性質
真核生物の遺伝子にはエキソンとイントロンが介在しており、遺伝子が転写されるときにはそのままコピーされ(mRNA前駆体)、mRNA前駆体からスプライシングとよばれる工程によってイントロンが除かれ完成型mRNAがつくられます。イントロンの塩基配列は、GUで始まりAGで終わることと、分岐部位とよばれるものが存在することが明らかになっています。イントロンは、分岐部位のアデニル酸残基の2’-OHがイントロンの5’末端のリン酸を求核攻撃し、これによって生じた上流エキソンの3’-OHが3’スプライス部位と結合することによって除去されます。イントロンが遊離する際、投げ縄構造(ラリアット構造)を形成します。スプライシングされていないmRNA前駆体は、ヘテロ核RNA(heterogeneous nuclear RNA; hnRNA)とよばれます。
活用例
cDNAのみを検出することが重要であるRT-PCRによる遺伝子発現解析実験において、ゲノムDNAがエキソン、イントロン構造を持っていることを利用して、ゲノムDNA由来の増幅が起こらないようなプライマーを設計することができます。まず、目的遺伝子のゲノム構造を確認し、サイズの大きなイントロンを選び出します。そして、このイントロンを挟む2つのエキソン上に上流プライマー、下流プライマーをそれぞれ設計します。イントロンのサイズが十分に大きければ、ゲノムDNA由来の増幅が起こりません。イントロンのサイズが小さい場合にも、ゲノム由来のPCR増幅産物は、mRNA由来のものよりもサイズが大きくなるため、融解曲線分析で区別できます。
歴史的背景
1977年に真核生物の遺伝子上にイントロンが発見されて以来、イントロンの起源・進化・機能に関しての論争が続いています。イントロンの出現時期に関して、二つの立場に分かれて活発な議論が展開されています。Gilbertらは、イントロンの前生説(introns early)を唱えました。その主張は、イントロンは原始的な生命において既に存在し、現在の古細菌や真正細菌、下等な真核生物のゲノムから失われた、というものでした。これに対して、Cavalier-Smithらは、イントロンの後生説(introns late)を唱え、イントロンは真核生物において出現し徐々に蓄積されてきたと主張します。これまで、様々なデータセットを用いて解析が行われてきましたが、いまだこの議論に決着はついていません。
お仕事をお探しの方へ
研究職のお仕事を探すならWDB
WDBの求人サイトでは「遺伝子解析」に関連する求人情報を多数掲載しています。
このページをシェアする
関連サイト
新卒学生の方はこちら


求職者の方はこちら


法人の方はこちら

